
Seguramente conoces el código genético, que recoge cómo a partir del ADN la información genética se expresa en forma de proteínas. ¿Sabías que éste no es el único código que existe en genética? Además de los ácidos nucleicos, las proteínas histonas también pueden codificar una determinada información y afectar a la expresión de los genes. Hablamos del código de las histonas.
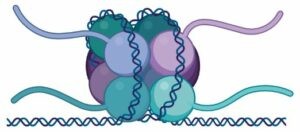
En eucariotas el ADN está empaquetado y compactado en el núcleo celular. Esto se consigue gracias a la interacción entre el ADN y varias proteínas que forman la cromatina, una especie de fibra que se condensa para formar los cromosomas. La unidad fundamental de la cromatina es el nucleosoma, que consiste en un octámero de proteínas llamadas histonas, el ADN enrollado a su alrededor y otra histona que lo sujeta.
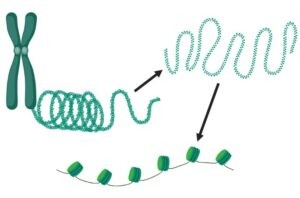
Formación de los nucleosomas: un elemento clave para el código de las histonas
Los nucleosomas se pueden ensamblar y desensamblar durante diferentes procesos celulares. Por ejemplo, para que pueda producirse la replicación del ADN las histonas deben separarse de él, para que la polimerasa y otras enzimas puedan acceder a todo el ADN.
En la formación del nucleosoma, primero tiene lugar el ensamblaje de dos histonas H3 y dos histonas H4 para formar el tetrámero central. El ADN (150 pares de bases aproximadamente) se enrolla alrededor de ese tetrámero y luego se añaden dos dímeros formados por las histonas H2A y H2B cada uno. Finalmente, se une una histona H1, que mantiene al ADN sujeto alrededor del octámero.
Hasta ahora hemos visto el papel estructural de las histonas en la cromatina, pero también juegan un importante papel en la expresión génica. Hay dos tipos de cromatina: la eucromatina, que es transcripcionalmente activa, y la heterocromatina, que es transcripcionalmente inactiva. La heterocromatina está más condensada que la eucromatina, por lo que es más fácil acceder a la eucromatina y poder transcribir esos genes. Las histonas participan en la determinación del tipo de estructura de la cromatina. Por lo tanto regulan la expresión génica.
Modificaciones del código de las histonas
Las histonas pueden sufrir varios tipos de modificaciones (marcas epigenéticas), cada una de las cuales tiene un efecto diferente. Esas marcas influyen en el estado de la cromatina y por tanto en la expresión de los genes. Hay una gran variedad de modificaciones y muchas combinaciones posibles, pues todas las histonas pueden modificarse. Esto es como un lenguaje, el código de las histonas.
En todo este proceso de modificación de las histonas participan distintas proteínas:
- Los lectores son las proteínas encargadas de reconocer las modificaciones. Leen la información y según ella llevan a los escritores y a los borradores donde sean necesarios.
- Los escritores son las proteínas escritoras. Son las que añaden modificaciones a las histonas, escriben la información, guiados por los lectores.
- Los borradores se encargan de eliminar las marcas epigenéticas, también guiados por los lectores.
Actualmente se ha descifrado el significado, es decir, los efectos sobre la expresión génica, de muchos tipos de modificaciones, pero todavía quedan algunas incógnitas sin resolver. Las modificaciones más conocidas son la acetilación, la metilación, la ubiquitinación y la fosforilación.
– Acetilación
La acetilación consiste en la adición de un grupo acilo al aminoácido lisina. El ADN tiene carga negativa debido a los grupos fosfato, por tanto se une a las histonas por la atracción entre cargas opuestas de las lisinas, que tienen carga positiva. El grupo acilo neutraliza la carga positiva de la lisina, por lo que se pierde afinidad entre el ADN y la histona con acetilaciones, lo que facilita la separación. Como consecuencia, hay una mayor accesibilidad para las enzimas implicadas en la transcripción. Así pues, la acetilación se asocia con regiones transcripcionalmente activas de la cromatina, como la eucromatina, pues promueve la transcripción de los genes.
– Metilación
La metilación es la adición de un grupo metilo a dos posibles aminoácidos: la lisina y la arginina. En este caso, el efecto depende de la posición y del tipo de aminoácidos en las histonas en los que tenga lugar la metilación. Cuando se metilan las argininas se activa la transcripción, mientras que cuando se metilan las lisinas el efecto puede ser tanto la activación como la represión de la transcripción. La metilación de las histonas está implicada, por ejemplo, en el proceso de respuesta a daño celular.
– Ubiquitinación
En la ubiquitinación se añade una proteína ubiquitina o una cadena de varias ubiquitinas a las histonas. Cuando se añade una sola ubiquitina (monoubiquitinación) los efectos pueden ser muy diversos. Sobre todo está implicado en la regulación de señalización celular. Por otro lado, cuando se añaden varias ubiquitinas (poliubiquitinación) se puede promover la degradación de histonas o se pueden reclutar distintas proteínas, que pueden realizar una amplia variedad de funciones.
– Fosforilación
En la fosforilación se añade un grupo fosfato a determinados aminoácidos de las histonas, más concretamente a aquellos que tengan grupos hidroxilo. Esta modificación está implicada sobre todo en la compactación de la cromatina. Se ha observado que esos grupos fosfato pueden reclutar distintas proteínas implicadas en dicho proceso.
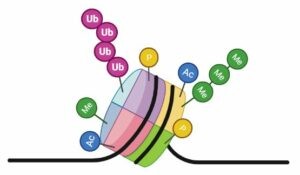
Cabe destacar que las modificaciones en las histonas no actúan por libre. El código de las histonas no es estanco para cada una. Existe “comunicación” entre las distintas modificaciones, haciendo que el código de las histonas sea muy complicado de descifrar. Los lectores pueden leer una determinada marca, y guiar a los escritores a que añadan otra distinta, que por su parte podría hacer que los borradores borren otra marca que había anteriormente, etc.
Finalmente, es importante quedarse con la idea de que, como las marcas epigenéticas en las histonas regulan la transcripción de los genes y tienen efectos a gran escala, pues afectan a muchos procesos fisiológicos. En algunas enfermedades, como el cáncer, se ha observado una desregulación en los procesos de modificación de histonas. Por tanto, es importante que las modificaciones sean leídas, borradas y escritas correctamente.
Fuentes:
Barnes CE, English DM, Cowley SM. Acetylation & Co: an expanding repertoire of histone acylations regulates chromatin and transcription. Essays Biochem. 2019 Apr 23;63(1):97-107. doi: 10.1042/EBC20180061. PMID: 30940741; PMCID: PMC6484784.
Gong F, Miller KM. Histone methylation and the DNA damage response. Mutat Res. 2019 Apr-Jun;780:37-47. doi: 10.1016/j.mrrev.2017.09.003. Epub 2017 Sep 23. PMID: 31395347; PMCID: PMC6690396.
Rothbart SB, Strahl BD. Interpreting the language of histone and DNA modifications. Biochim Biophys Acta. 2014 Aug;1839(8):627-43. doi: 10.1016/j.bbagrm.2014.03.001. Epub 2014 Mar 12. PMID: 24631868; PMCID: PMC4099259. https://pubmed.ncbi.nlm.nih.gov/24631868/
Zhang T, Cooper S, Brockdorff N. The interplay of histone modifications – writers that read. EMBO Rep. 2015 Nov;16(11):1467-81. doi: 10.15252/embr.201540945. Epub 2015 Oct 15. PMID: 26474904; PMCID: PMC4641500.
Si te ha gustado esta entrada y te interesa la genética, descubre nuestros nuevos cursos y formación universitaria.




