EN BUSCA DEL ORGANISMO MÁS ANTIGUO: EVIDENCIA INDIRECTA
Hasta ahora hemos intentado comprender cómo surgió la vida a partir del estudio de la química prebiótica y de las condiciones de la Tierra primigenia. A esta aproximación se la conoce como Bottom up, de abajo a arriba. En este apartado vamos a explicar la aproximación complementaria, llamada Top down, de arriba abajo. Mediante esta aproximación los científicos intentan establecer el árbol filogenético (como un árbol genealógico de distintas especies) de toda la historia de la vida, para así intentar explicar cómo fue el organismo más antiguo (Ikehara, 2016).
Charles Darwin, en 1856, determina en el “Origen de las Especies” que unas especies surgen mediante selección natural a partir de otras anteriores. Pero fue más allá. Desarrollando este argumento al máximo, Darwin escribe en el mismo libro: “Probablemente todos los seres orgánicos que alguna vez han vivido en esta Tierra han descendido de una forma de vida primordial” (Darwin C, 1859/2009). Pese a que Darwin no tenía ninguna prueba, dio en el clavo.
Antes de que la tecnología de secuenciación de genomas completos estuviera desarrollada, los biólogos y taxonomistas dividían la vida en la Tierra en 5 reinos: animales, plantas, hongos, moneras y protoctistas. Las moneras serían todos los seres unicelulares procariotas (sin núcleo) y los protoctistas todos los unicelulares eucariotas (con núcleo) (Woese, Kandler and Wheelis, 1990; Margulis, 1992; CAVALIER-SMITH, 1998).
Los árboles filogenéticos modernos dividen la vida en la Tierra de una manera muy distinta. Para determinar qué distancia evolutiva existe entre dos especies se determina el número de cambios en la secuencia de nucleótidos del genoma de ambas especies. Se asume que la tasa de variación en los genomas es constante en el tiempo. Esta característica denominada “reloj molecular” ha sido desarrollada hasta ofrecer resultados muy fiables. Por ejemplo, si en una secuencia de nucleótidos de una especie una T cambia a una A, tras unos millones de años la A cambia a G, y tras otro millón de años la G cambia a una T, detectaríamos que en esa posición la letra no ha cambiado. Mediante desarrollo de modelos matemáticos este tipo de sucesos se tienen en cuenta (Kumar, 2005).
Cuando comparamos las secuencias de todos los seres vivientes (cuyo genoma se ha secuenciado) obtenemos algo así.
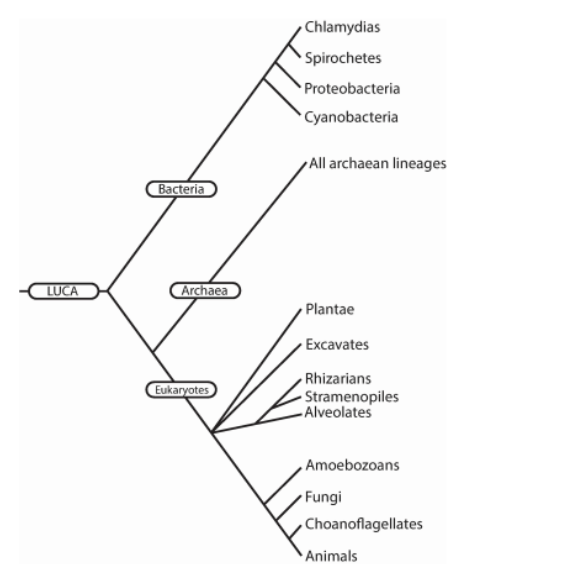
La vida se divide en 3 dominios: bacterias, arqueas y eucariotas. Las bacterias y arqueas representan el mundo procariota (todos son unicelulares), células pequeñas y relativamente simples sin núcleo. Los eucariotas se dividen en organismos unicelulares (protoctistas) y pluricelulares, dando lugar a toda la flora y fauna que conocemos, incluidos nosotros (Woese, Kandler and Wheelis, 1990).
Al analizar los árboles filogenéticos, obtenemos bifurcaciones. Éstas, hacen referencia a eventos de especiación, en el que una especie da lugar a dos especies distintas. De modo que los nodos (justo el punto en el que hay una bifurcación) representa al antepasado común de las dos ramas (especies) a las que dio lugar. Si analizamos el árbol, hubo un ancestro que dio lugar a los animales y a los hongos, más atrás en el tiempo, compartimos un antepasado con las plantas, más atrás aún las arqueas y los eucariotas compartieron un antepasado común, y más atrás aún ese antepasado y las bacterias compartieron un antepasado, el LUCA. LUCA son las siglas en inglés de Last Universal Common Ancestor (Último Antepasado Común Universal). Es decir, toda forma de vida en la Tierra surgió de una única especie. Los científicos intentan comprender cómo era LUCA para acercarnos un poco más al origen de la vida (Mayr, 1974; Penny, Hendy and Steel, 1992; Ciccarelli et al., 2006; Di Giulio, 2007; Podani, 2019).
Sin embargo, LUCA, aunque es nuestro primer ancestro, no es, ni de lejos, el primer organismo vivo. Muchas formas de vida antes de LUCA debieron surgir de forma paralela. Se cree que LUCA ya tenía genoma de ADN, además de que codificaba proteínas con mensaje genético, por lo que probablemente tenía ribosoma. Es decir, no se diferenciaba mucho de una bacteria actual. Los científicos estiman que LUCA vivió hace unos 3500 millones de años (Weiss et al., 2016; Eme et al., 2017).
Aunque sabemos que LUCA existió, es mediante evidencia genómica, pero no directa. Al comparar todas las secuencias de especies vivientes y extintas, los resultados indican que hubo un ancestro de todas ellas. Sin embargo, no todas las secuencias de ADN presentes en el genoma de una especie se encuentran ahí por herencia vertical, es decir, la información genética que una célula madre le transfiere a una célula hija, y por ello no son susceptibles de ser analizadas mediante un árbol filogenético.
Un fenómeno muy importante en la evolución es la transferencia horizontal de genes (THG) (Keeling and Palmer, 2008; Fournier, Andam and Gogarten, 2015). Ésta se da principalmente mediante los virus. Ciertos tipos de virus, cuando infectan una célula, insertan su genoma dentro del genoma de la célula hospedadora. Así, cuando la célula replique su genoma, también replicará el genoma del virus, sin que éste tenga que hacer nada. Cuando se dan las condiciones propicias, la secuencia vírica se activa y se escinde del genoma, para formar nuevas partículas víricas e infectar nuevas células. En el proceso de escisión, es frecuente que un virus “arranque” un trozo de secuencia del genoma de la célula hospedadora y se lo lleve consigo (Keeling and Palmer, 2008; Callier, 2019)(este mecanismo se cree que causa hoy en día hasta un 20% de los tumores (Liao, 2006; Martin and Gutkind, 2008)). Cuando ese virus infecte a una nueva célula, le introducirá la porción del genoma, y los genes que pueda albergar, en el genoma de la nueva célula. De esta manera se ha dado un evento de THG.
Además, también es posible que por accidente el genoma vírico se dañe y quede atrapado en el genoma de la célula que ha infectado, formando así para siempre parte de su genoma (y del de su estirpe).
Estos eventos de THG se consideran muy importantes en la historia de la evolución de la vida, y sobre todo de la vida temprana, en la que el número de genes en un individuo sería muy reducido. Este concepto de transferencia horizontal fue llevado al extremo por Lynn Margulis al proponer que las células eucariotas actuales surgieron de una fagocitosis entre arqueas y bacterias, es decir, que una célula de tipo arquea se “comió” a una bacteria, pero no la digirió, generando así la mitocondria, un orgánulo de las células eucariotas. Llamó a esta teoría endosimbiosis, y hoy en día tiene un gran fundamento experimental. Por ejemplo, las mitocondrias tienen su propio genoma, que, además, es de tipo bacteriano, muy distinto al genoma nuclear. (Margulis, 1992; Di Giulio, 2007; Wächtershäuser, 2007; Keeling and Palmer, 2008; Fournier, Andam and Gogarten, 2015; Eme et al., 2017; Moelling and Broecker, 2021).
No podemos saber cuándo se dieron los THG, por lo que no podemos reconstruir el árbol de la vida a partir de genomas completos. En su lugar, debe recurrirse a genes específicos o grupos de genes presentes en todos los seres vivos. Los más usados son los genes que codifican los ARNs ribosómicos (Syvanen, 2002; Peplies et al., 2008; Yarza et al., 2008; Tsukuda, Kitahara and Miyazaki, 2017).
Otra consecuencia de los eventos THG es que quizás no existe un LUCA, sino muchos. Según este enfoque, a partir de una población heterogénea de LUCAS, se irían compartiendo genes mediante THG entre ellos, lo que iría generando una mayor biodiversidad. Por lo que, pese a todo, quizás no tengamos un único ancestro común sino varios (Woese, 1998).
EN BUSCA DEL ORGANISMO MÁS ANTIGUO: EVIDENCIA DIRECTA
Pero ¿cuál es el primer fósil del que se tiene constancia? En las costas de Australia se encontró en 2016 estromatolitos de 3400 millones de años (Duda et al., 2016). Los estromatolitos son estructuras minerales que se generan cuando bacterias fotosintéticas captan el CO2 del aire y lo fijan biológicamente formando carbonatos.
El primer fósil del que se tiene registro es una bacteria filamentosa con una antigüedad de 3465 millones de años.
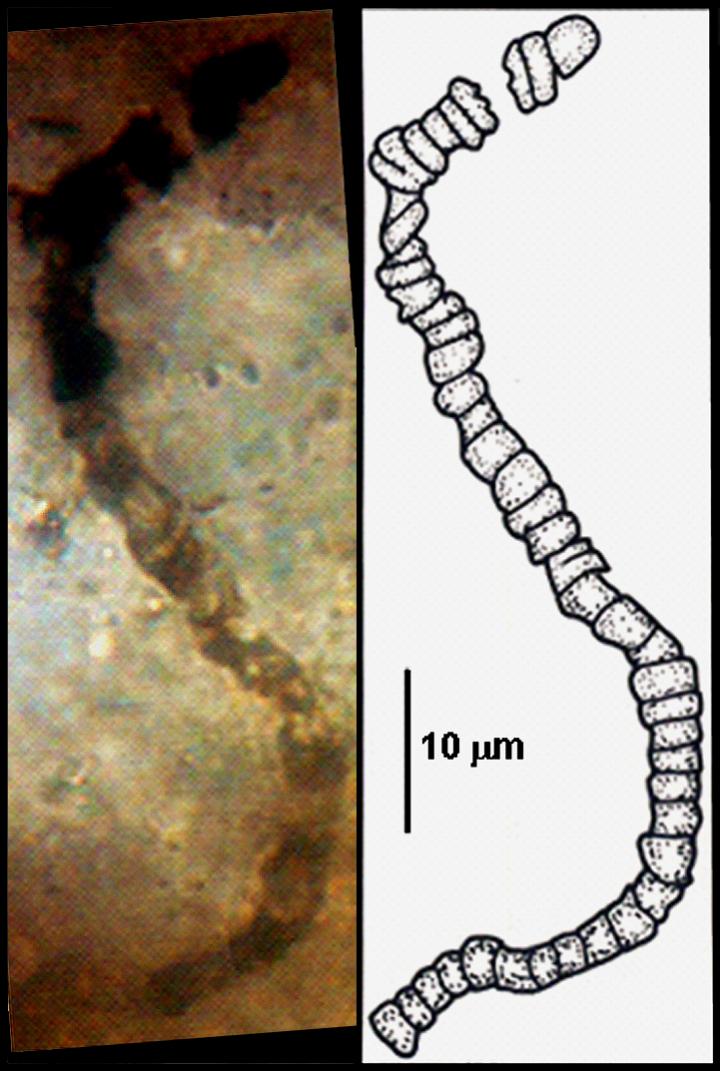
Ha habido descripciones de posibles formas de vida mucho más antiguas, pero únicamente basadas en la morfología. El científico español Juan Manuel Garcia Ruiz es famoso mundialmente por la descripción de los biomorfos. Éstos son estructuras complejas autoensambladas con una morfología muy similar a algunos tipos de seres unicelulares pero que están constituidos únicamente de materia inorgánica (Kellermeier et al., 2013; García-Ruiz, van Zuilen and Bach, 2020b, 2020a; Kotopoulou et al., 2021).
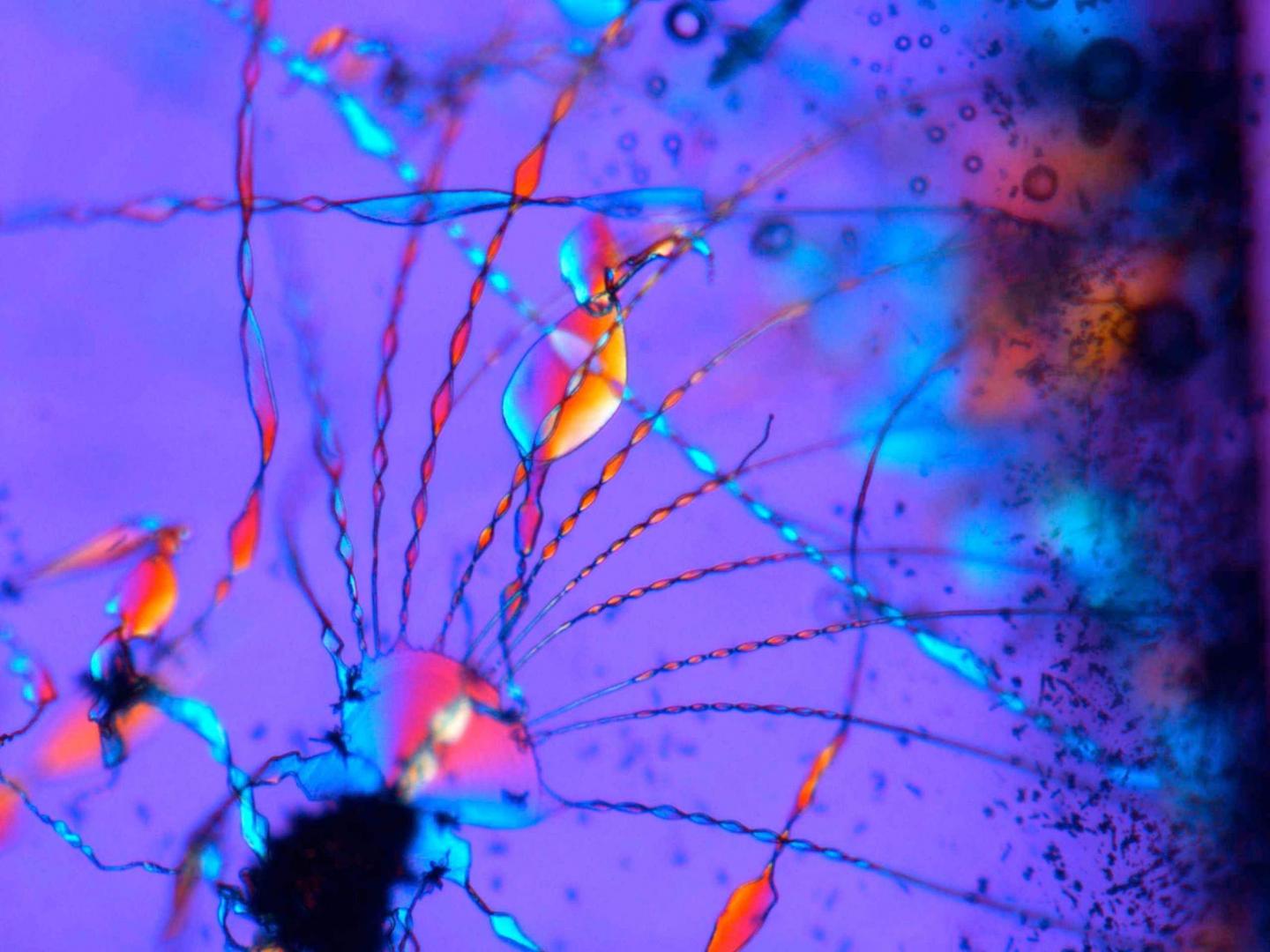
Teniendo en cuenta estas estructuras se debe poner en tela de juicio cualquier descubrimiento de formas de vida terrestres o extraterrestres basadas únicamente en la morfología.
De modo que, si no podemos confiar en la morfología, sólo nos queda la química. Cada elemento químico está formado en su núcleo por un determinado número de protones y neutrones. El número de protones determina qué elemento es: si tiene 6 protones es carbono, si tiene 14 es silicio. El número de neutrones, en cambio, puede variar en un mismo elemento. Los distintos átomos de carbono, por ejemplo, con distinto número de neutrones en su núcleo, se llaman isótopos del carbono. La corteza terrestre y cualquier formación mineral tiene unas determinadas proporciones de los isótopos de cada elemento. La vida, sin embargo, tiene preferencia por utilizar un determinado tipo de isótopo, en concreto, la vida prefiere el isótopo 12 del carbono al isótopo 13 o 14 (Zubarev, 2011). Si un mineral está enriquecido en isótopo 12, significa que probablemente haya sido fijado (formado) por un ser vivo. Este tipo de evidencia de metabolismo, llamados fósiles químicos, son las primeras evidencias de vida en la Tierra.
Las señales de carbono fijado biológicamente datan de 4,1 miles de millones de años en los casos más dudosos y de 3,8 en los más aceptados. De modo que después del gran bombardeo tardío comentado anteriormente, la vida pudo haber necesitado como máximo unos 350 millones de años en desarrollarse, como mínimo unos 50 millones (Bell et al., 2015; Dodd et al., 2017; Hassenkam et al., 2017; Betts et al., 2019).
¿EXISTE UN DETERMINISMO EN LA APARICIÓN DE VIDA?
Aunque pueda parecer mucho tiempo, 50 millones de años en escalas geológicas no lo es en absoluto. De hecho, los expertos no se ponen de acuerdo en si es suficiente tiempo para que se forme la vida. Por ello, se han desarrollado dos posiciones teóricas contrarias. La primera postura asume que si la vida se generó en la Tierra, el hecho de que pasase tan poco tiempo hasta desarrollarse indicaría que la vida surge de la química en cuanto puede: hay una tendencia a la formación de vida en la química. De esta postura se deduce que el Universo estaría lleno de vida si se dan las condiciones apropiadas.
Por otro lado, si la formación de vida en la Tierra se debió a un cúmulo inimaginable de casualidades y condiciones altamente improbables, la formación de vida sería un fenómeno extremadamente improbable, y probablemente estaríamos solos en la galaxia.
Estas dos posturas, tan extremas y opuestas son hoy en día todavía objeto de debate, y los científicos no saben hacia cual inclinarse (Monod, J 1970/2016; Aguilera, JA 2017; Briones, C 2020).
Referencias
Aguilera, JA. El Origen de La Vida. RBA Editorial. 2019. ISBN: 9788491873341.
Alberty, R. A. (2006) ‘Relations between biochemical thermodynamics and biochemical kinetics’, Biophysical Chemistry, 124(1), pp. 11–17. doi: 10.1016/j.bpc.2006.05.024.
Andrulis, E. D. (2011a) ‘Theory of the origin, evolution, and nature of life’, Life. Life (Basel), 2(1), pp. 1–105. doi: 10.3390/life2010001.
Arney, G., et al. (2018) ‘Venus as an analog for hot earths’, arXiv. doi: 10.2458/azu_uapress_9780816540068-ch014.
Ballen, C. J. and Greene, H. W. (2017) ‘Walking and talking the tree of life: Why and how to teach about biodiversity’, PLoS Biology, 15(3), pp. 1–11. doi: 10.1371/journal.pbio.2001630.
Bang, B. and Bang, B. (2008) ‘Wilkinson Microwave Anisotropy Probe Cosmology: The Study of the Universe’, pp. 10–19.
Bell, E. A. et al. (2015) ‘Potentially biogenic carbon preserved in a 4.1 billion-year-old zircon’, Proceedings of the National Academy of Sciences of the United States of America, 112(47), pp. 14518–14521. doi: 10.1073/pnas.1517557112.
Betts, H. C. et al. (2019) ‘Europe PMC Funders Group Integrated genomic and fossil evidence illuminates life ’ s early evolution and eukaryote origins’, 2018(10), pp. 1556–1562. doi: 10.1038/s41559-018-0644-x.Integrated.
Briones, C. ¿Estamos solos? En busca de otras vidas en el cosmos. CRITICA. 2020. ISBN: 9788491992219.
Callier, V. (2019) ‘Gene transfers from bacteria and viruses may be shaping complex organisms’, Proceedings of the National Academy of Sciences of the United States of America, 116(28), pp. 13714–13716. doi: 10.1073/pnas.1909030116.
CAVALIER-SMITH, T. (1998) ‘A revised six-kingdom system of life’, Biological Reviews of the Cambridge Philosophical Society, 73(3), pp. 203–266. doi: 10.1017/s0006323198005167.
Ciccarelli, F. D. et al. (2006) ‘of a Highly Resolved Tree of Life’, Science, 311(March), pp. 1283–1287. Available at: http://www.ncbi.nlm.nih.gov/pubmed/16513982.
Cleaves, H. J. et al. (2012) ‘Mineral–organic interfacial processes: Potential roles in the origins of life’, Chemical Society Reviews, 41(16), pp. 5502–5525. doi: 10.1039/c2cs35112a.
Cleaves, H. J. (2013) ‘Prebiotic chemistry: Geochemical context and reaction screening’, Life, 3(2), pp. 331–345. doi: 10.3390/life3020331.
Darwin, CR. El Origen De Las Especies Por Medio De La Selección Natural. 2009. Alianza editorial. ISBN: 978-84-206-6867-3 (Obra original publicada en: 1858).
Deamer, D. et al. (2002) ‘ASTROBIOLOGY Volume 2, Number 4, 2002 © Mary Ann Liebert, Inc.’, Astrobiology, 2(4), pp. 371–381.
Deamer, D. (2017) ‘The role of lipid membranes in life’s origin’, Life, 7(1). doi: 10.3390/life7010005.
Deamer, D. and Weber, A. L. (2010) ‘Bioenergetics and Life ’ s Origins’, pp. 1–16.
Dodd, M. S. et al. (2017) ‘Evidence for early life in Earth’s oldest hydrothermal vent precipitates’, Nature. Nature Publishing Group, 543(7643), pp. 60–64. doi: 10.1038/nature21377.
Duda, J. P. et al. (2016) ‘A rare glimpse of paleoarchean life: Geobiology of an exceptionally preserved microbial mat facies from the 3.4 ga strelley pool formation, Western Australia’, PLoS ONE, 11(1), pp. 1–18. doi: 10.1371/journal.pone.0147629.
Dwyer, D. S. (2008) ‘Amino Acids: Chemical Properties’, Wiley Encyclopedia of Chemical Biology, (July). doi: 10.1002/9780470048672.wecb007.
Eme, L. et al. (2017) ‘Archaea and the origin of eukaryotes’, Nature Reviews Microbiology, 15(12), pp. 711–723. doi: 10.1038/nrmicro.2017.133.
Fournier, G. P., et al. (2015) ‘Ancient horizontal gene transfer and the last common ancestors Phylogenetics and phylogeography’, BMC Evolutionary Biology. ???, 15(1), pp. 1–18. doi: 10.1186/s12862-015-0350-0.
García-Ruiz, J. M., et al. (2020a) ‘Mineral self-organization on a lifeless planet’, Physics of Life Reviews. Elsevier B.V., 34–35, pp. 62–82. doi: 10.1016/j.plrev.2020.01.001.
Di Giulio, M. (2007) ‘The universal ancestor and the ancestors of Archaea and Bacteria were anaerobes whereas the ancestor of the Eukarya domain was an aerobe’, Journal of Evolutionary Biology, 20(2), pp. 543–548. doi: 10.1111/j.1420-9101.2006.01259.x.
Gosavi, S. (2013) ‘Understanding the Folding-Function Tradeoff in Proteins’, PLoS ONE, 8(4). doi: 10.1371/journal.pone.0061222.
Gowanlock, M. G., Patton, D. R. and McConnell, S. M. (2011) ‘A model of habitability within the Milky Way galaxy’, Astrobiology, 11(9), pp. 855–873. doi: 10.1089/ast.2010.0555.
Halliday, A. N. and Able, T. (2006) ‘The Origin of the Earth What ’ s New ? FORMATION’, Earth, pp. 205–210.
Hassenkam, T. et al. (2017) ‘Elements of Eoarchean life trapped in mineral inclusions’, Nature. Nature Publishing Group, 548(7665), pp. 78–81. doi: 10.1038/nature23261.
Hazen, R. M. and Sverjensky, D. A. (2010) ‘Mineral surfaces, geochemical complexities, and the origins of life.’, Cold Spring Harbor perspectives in biology, 2(5), pp. 1–21. doi: 10.1101/cshperspect.a002162.
Higgs, P. G. and Lehman, N. (2015) ‘The RNA World: Molecular cooperation at the origins of life’, Nature Reviews Genetics. Nature Publishing Group, 16(1), pp. 7–17. doi: 10.1038/nrg3841.
Ikehara, K. (2016) ‘Evolutionary steps in the emergence of life deduced from the bottom-up approach and GADV hypothesis (Top-down approach)’, Life, 6(1), pp. 1–15. doi: 10.3390/life6010006.
Jadhav, V. R. and Yarus, M. (2002) ‘Coenzymes as coribozymes’, Biochimie, 84(9), pp. 877–888. doi: 10.1016/S0300-9084(02)01404-9.
Johnson, J. A., et al. (2020) ‘The origin of the elements: A century of progress’, Philosophical Transactions of the Royal Society A: Mathematical, Physical and Engineering Sciences, 378(2180). doi: 10.1098/rsta.2019.0301rsta20190301.
Keeling, P. J. and Palmer, J. D. (2008) ‘Horizontal gene transfer in eukaryotic evolution’, Nature Reviews Genetics, 9(8), pp. 605–618. doi: 10.1038/nrg2386.
Kellermeier, M. et al. (2013) Experimental techniques for the growth and characterization of silica biomorphs and silica gardens, Methods in Enzymology. doi: 10.1016/B978-0-12-416617-2.00011-4.
Kellermeier, M., Cölfen, H. and García-Ruiz, J. M. (2012) ‘Silica biomorphs: Complex biomimetic hybrid materials from “sand and chalk”’, European Journal of Inorganic Chemistry, (32), pp. 5123–5144. doi: 10.1002/ejic.201201029.
Khurana, K. K., et al. (2002) ‘Using Surface Observatories’, Astrobiology, 2(1), pp. 93–103.
Kitadai, N. and Maruyama, S. (2018) ‘Origins of building blocks of life: A review’, Geoscience Frontiers. Elsevier, 9(4), pp. 1117–1153. doi: 10.1016/j.gsf.2017.07.007.
Kotopoulou, E. et al. (2021) ‘Nanoscale Anatomy of Iron-Silica Self-Organized Membranes: Implications for Prebiotic Chemistry’, Angewandte Chemie – International Edition, 60(3), pp. 1396–1402. doi: 10.1002/anie.202012059.
Kumar, S. (2005) ‘Molecular clocks: Four decades of evolution’, Nature Reviews Genetics, 6(8), pp. 654–662. doi: 10.1038/nrg1659.
Lambert, J. F. (2008) ‘Adsorption and polymerization of amino acids on mineral surfaces: A review’, Origins of Life and Evolution of Biospheres, 38(3), pp. 211–242. doi: 10.1007/s11084-008-9128-3.
Lauro, S. E. et al. (2021) ‘Multiple subglacial water bodies below the south pole of Mars unveiled by new MARSIS data’, Nature Astronomy. Springer US, 5(1), pp. 63–70. doi: 10.1038/s41550-020-1200-6.
Lazcano, A. and Miller, S. L. (1996) ‘The origin and early evolution of life: Prebiotic chemistry, the pre-RNA world, and time’, Cell, 85(6), pp. 793–798. doi: 10.1016/S0092-8674(00)81263-5.
Liao, J. B. (2006) ‘Cancer Issue: Viruses and Human Cancer’, The Yale Journal of Biology and Medicine, 79(3–4), p. 115. Available at: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC1994798/.
Lineweaver, C. H. (2001) ‘An Estimate of the Age Distribution of Terrestrial Planets in the Universe: Quantifying Metallicity as a Selection Effect’, Icarus, 151(2), pp. 307–313. doi: 10.1006/icar.2001.6607.
Liu, H. et al. (2016) ‘A Facile Chemical-Free and Universal Method for Transfer of Ultrathin Graphene-Based Films’, Advanced Materials Interfaces, 3(18). doi: 10.1002/admi.201600540.
Margulis, L. (1992) ‘Biodiversity: molecular biological domains, symbiosis and kingdom origins’, BioSystems, 27(1), pp. 39–51. doi: 10.1016/0303-2647(92)90045-Z.
Martin, D. and Gutkind, J. S. (2008) ‘Human tumor-associated viruses and new insights into the molecular mechanisms of cancer’, Oncogene. Nature Publishing Group, 27, pp. S31–S42. doi: 10.1038/onc.2009.351.
Mayr, E. (1974) ‘Cladistic analysis or cladistic classification?’, Journal of Zoological Systematics and Evolutionary Research, 12(1), pp. 94–128. doi: 10.1111/j.1439-0469.1974.tb00160.x.
McIntyre, S. R. N., et al. (2019) ‘Planetary magnetism as a parameter in exoplanet habitability’, Monthly Notices of the Royal Astronomical Society. Oxford University Press, 485(3), pp. 3999–4012. doi: 10.1093/mnras/stz667.
Mcnichol, J. C. et al. (2012) ‘Jesse C. McNichol’. doi: 10.1007/978-94-007-2941-4.
Moelling, K. and Broecker, F. (2021) ‘Viroids and the origin of life’, International Journal of Molecular Sciences, 22(7), pp. 1–13. doi: 10.3390/ijms22073476.
Monod, J. El Azar y La Necesidad. 2016. TUSQUETS EDITORES. ISBN: 9788490662120 (Obra original publicada en: 1970).
Montmerle, T. et al. (2006) ‘Solar system formation and early evolution: The first 100 million years’, Earth, Moon and Planets, 98(1–4), pp. 39–95. doi: 10.1007/s11038-006-9087-5.
Montoya-Herrera, F. L. (1988) ‘No Titleآبهای زیرزمینی’, آب و خاک, 19(31), pp. 21–38.
n, Eunsung Mouradian, M. M. (2008) ‘基因的改变NIH Public Access’, Bone, 23(1), pp. 1–7. doi: 10.1016/j.tibs.2009.01.005.Co-evolution.
News, F. (2018) ‘Cataclysm’s End’, Nature News. Available at: https://www.nature.com/magazine-assets/d41586-018-01074-6/d41586-018-01074-6.pdf.
Penny, D., Hendy, M. D. and Steel, M. A. (1992) ‘Progress with methods for constructing evolutionary trees’, Trends in Ecology and Evolution, 7(3), pp. 73–79. doi: 10.1016/0169-5347(92)90244-6.
Peplies, J. et al. (2008) ‘A standard operating procedure for phylogenetic inference (SOPPI) using (rRNA) marker genes’, Systematic and Applied Microbiology, 31(4), pp. 251–257. doi: 10.1016/j.syapm.2008.08.003.
Podani, J. (2019) ‘The Coral of Life’, Evolutionary Biology. Springer US, 46(2), pp. 123–144. doi: 10.1007/s11692-019-09474-w.
Saladino, R. et al. (2018) ‘Rewarming the Primordial Soup: Revisitations and Rediscoveries in Prebiotic Chemistry’, ChemBioChem, 19(1), pp. 22–25. doi: 10.1002/cbic.201700534.
Scheller, E. L. et al. (2021) ‘Long-term drying of Mars by sequestration of ocean-scale volumes of water in the crust’, Science, 372(6537), pp. 56–62. doi: 10.1126/science.abc7717.
Schopf, J. W. (1993) ‘Microfossils of the early Archean apex chert: New evidence of the antiquity of life’, Science, 260(5108), pp. 640–646. doi: 10.1126/science.260.5108.640.
Spalding, C. and Fischer, W. W. (2019) ‘A shorter Archean day-length biases interpretations of the early Earth’s climate’, Earth and Planetary Science Letters, 514, pp. 28–36. doi: 10.1016/j.epsl.2019.02.032.
Stanley L. Miller (1953) ‘Miller-Science-1953.pdf’, Science, pp. 528, 529. Available at: http://www.chm.bris.ac.uk/motm/glycine/Miller-Science-1953.pdf.
Syvanen, M. (2002) ‘Rates of ribosomal RNA evolution are uniquely accelerated in Eukaryotes’, Journal of Molecular Evolution, 55(1), pp. 85–91. doi: 10.1007/s00239-001-2306-9.
Tsukuda, M., et al. (2017) ‘Comparative RNA function analysis reveals high functional similarity between distantly related bacterial 16 S rRNAs’, Scientific Reports. Springer US, 7(1), pp. 1–8. doi: 10.1038/s41598-017-10214-3.
Wächtershäuser, G. (2007) ‘On the chemistry and evolution of the pioneer organism’, Chemistry and Biodiversity, 4(4), pp. 584–602. doi: 10.1002/cbdv.200790052.
Wagner, K. et al. (2021) ‘Imaging low-mass planets within the habitable zone of α Centauri’, Nature Communications, 12(1), pp. 1–7. doi: 10.1038/s41467-021-21176-6.
Waller, W. H. (2019) ‘Life in the Milky Way’, The Milky Way, 303(January), pp. 251–276. doi: 10.2307/j.ctt28556v.16.
Weiss, M. C. et al. (2016) ‘The physiology and habitat of the last universal common ancestor’, Nature Microbiology. Nature Publishing Group, 1(9), pp. 1–8. doi: 10.1038/nmicrobiol.2016.116.
Woese, C. (1998) ‘The universal ancestor’, Proceedings of the National Academy of Sciences of the United States of America, 95(12), pp. 6854–6859. doi: 10.1073/pnas.95.12.6854.
Woese, C. R., et al. (1990) ‘Towards a natural system of organisms: Proposal for the domains Archaea, Bacteria, and Eucarya’, Proceedings of the National Academy of Sciences of the United States of America, 87(12), pp. 4576–4579. doi: 10.1073/pnas.87.12.4576.
Yarza, P. et al. (2008) ‘The All-Species Living Tree project: A 16S rRNA-based phylogenetic tree of all sequenced type strains’, Systematic and Applied Microbiology, 31(4), pp. 241–250. doi: 10.1016/j.syapm.2008.07.001.
Zahnle, K., Schaefer, L. and Fegley, B. (2010) ‘Earth’s earliest atmospheres.’, Cold Spring Harbor perspectives in biology, 2(10), pp. 1–17. doi: 10.1101/cshperspect.a004895.
Zubarev, R. A. (2011) ‘Role of stable isotopes in life-testing isotopic resonance hypothesis’, Genomics, Proteomics and Bioinformatics. Genomics, Proteomics & Bioinformatics, 9(1–2), pp. 15–20. doi: 10.1016/S1672-0229(11)60003-X.